Home 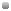 Генетика собак
Генетика собак
| Генетика собак - ПРИЗНАКИ, ОГРАНИЧЕННЫЕ ПОЛОМ |  |
 |
 |
|
Страница 4 из 65
Некоторые признаки, называемые зависимыми от пола, определяются генами, находящимися не в Х-хромосомах, а в одной из пар аутосом. Однако проявление таких признаков зависит от пола животного (независимо от сцепления с полом), что обычно связано с действием мужских или женских половых гормонов (в рецессивном состоянии признак отсутствует независимо от пола). Типичными примерами являются все вторичные половые признаки — комолость у некоторых пород овец, раннее облысение у людей, яйценоскость кур и наследование окраса у эр- ширских молочных коров, как показано в таблице 4. ТАБЛИЦА 4 Зависимость окраса эрширских молочных коров от пола Генотип Фенотип Быки Коровы ММ Махагон Махагон Mm Махагон Красный mm Красный Красный Некоторые признаки, ограниченные полом, проявляются только у самок, как, например, молочность, или у самцов, как крипторхизм, хотя носителями их могут быть и те и другие (так как они полигенные, а контролирующие их гены расположены в любых парах аутосом). Это осложняет генетические исследования и в некоторых случаях означает, что при селекции следует руководствоваться качеством потомства в целом, а не отдельно взятого животного, так как по одному полу невозможно сделать правильные выводы. Некоторые признаки, зависящие от пола, проявляются и у самцов, и у самок, но у одних из них они встречаются чаще, а у других — реже. Так, заячья губа и подагра, встречающиеся у людей, чаще всего наблюдаются все-таки у мужчин, а вот spina bifida (расщелина позвоночника — врожденная аномалия, характеризующаяся неполным закрытием позвоночного канала из-за отсутствия дуги нескольких позвонков, главным образом в поясничной области) и дисплазия тазобедренного сустава — у женщин. Наследование ограниченных полом признаков осуществляется в соответствии с законами Менделя. НЕПОЛНАЯ ПЕНЕТРАНТНОСТЬ При обсуждении доминантных и рецессивных аллелей предполагалось, что конкретный признак проявляется у животного в присутствии одного-единственного доминантного аллеля. Рецессивный же аллель для его проявления должен быть в гомозиготном состоянии. В этом случае мы имеем дело с так называемой полной (абсолютной) пенетрантностью {от лат. penetrare — проникновение). В некоторых же случаях, чаще всего у гетерозиготных особей (Аа), доминантный ген А в 75% случаев может вести себя как доминантный, а в 25% случаев — как рецессивный. В этом случае речь идет о неполной, а 75%-ной пенетран- тности. Причиной тому чаще всего служит полигенное наследование признака. При неполной пенетрантности заводчик сталкивается с определенными трудностями, ибо на первый взгляд результаты вязок не соответствуют законам Менделя, что связано с недостаточным количеством потомства для его анализа. ФЕНОКОПИИ Заводчики часто ошибочно полагают, что наследственность зависит только от генотипа, забывая о факторах индивидуального развития организма и влияния внешней среды. Так, травма суки во время щенности может значительно повлиять на развитие эмбриона, но не на его генотип, в отличие от внешних воздействий, способных вызвать мутации у плода. Особь с измененным генотипом называется фенокопией. Пожалуй, самым трагичным примером фенокопий у человека можно назвать случаи, связанные с применением беременными женщинами в 60-х годах талидомида — транквилизатора с тератогенным действием, приводящего к фокомелии или ахе- роподии у новорожденных (укорочению или отсутствию конечностей). Примером фенокопии служат и мягкие хрящи у собак со стоячими в норме ушами. Конечно, их можно поставить за счет фиксации ушных раковин, но такие собаки стойко будут передавать полустоячие уши своему потомству. ВАРИАЦИИ ЧИСЛА ПОЛОВЫХ ХРОМОСОМ Я уже писал, что в норме кариотип собак включает 78 хромосом, образующих 39 пар, включая одну половую: XX — у сук и XY — у кобелей. При изучении генного баланса, генетики обнаружили, что, даже если он и нормальный, в нем все равно встречаются отклонения. Вариации числа половых хромосом могут быть связаны с целым набором хромосом (эуплоидия) или с одной хромосомой (анэуплоидия). Пожалуй, самым известным примером такого явления служит синдром Дауна у человека, когда в двадцать первой паре хромосом (в норме их 23) появляется лишняя хромосома. Это происходит на определенной стадии формирования гамет и чаще наблюдается среди старородящих, как правило сорокалетних матерей. Возможно, подобные хромосомные аберрации происходят чаще, чем нам кажется, но мы не так часто видим их проявле- Рис. 12. Типы нехваток хромосом 1 — концевая (терминальная, или дефишиенси); 2 — две концевые нехватки, приводящие к образованию кольца; 3 — внутрихромосомная нехватка (делеция) ние из-за гибели гамет или ранней смертности плода. Болезнь Дауна (одна из форм олигофрении) — один из тех случаев, когда аберрация не настолько серьезная с генетической точки зрения, чтобы вызвать раннюю смерть эмбриона. А так как подобные аномалии встречаются в собаководстве чрезвычайно редко, мало кто из ученых придает им большое значение. Второй тип — дупликация (удвоение) хромосом. В этом случае одновременно с возникновением нехватки в результате разрыва хромосомы и потери участка внутри одной хромосомы могут накапливаться идентичные участки, несущие одни и те же гены. При дупликации происходит фенотипическое изменение признаков. Если в норме каждый участок хромосомы представлен одной дозой гена, то при удвоении или утроении несущего эти гены участка доза гена увеличивается в два или три раза. То есть у сук имеется не диплоидный набор (2Х:2А, то есть половой индекс, или соотношение между Х-хромосомами и аутосомами, равен единице), а три половые хромосомы — XXX, из-за чего они называются сверхсуками (они могут быть диплоидными — ЗХ:2А с половым индексом 1,5 или триплоидными — ЗХ:ЗА). У таких сук наблюдается нарушение астральных циклов (как правило, они стерильные), а если и нет, то при их вязке с нормальными кобелями рождаются интерсексы (2Х:ЗА с половым индексом 0,66) мужского или женского типов — гермафродиты. У кобелей комплекс XXY (синдром Клайфельтера) также сопровождается явлениями инфантилизма из-за недоразвития семенников и азооспермии. Лишняя Х-хромосома у котов обусловливает появление в помете самок черепахового окраса, реже самцов с половым набором XXY, страдающих стерилитетом. Животные с лишней половой хромосомой, как правило, стерильные. Типичный пример — мул (гибрид кобылы, имеющей 64 хромосомы, и осла, у которого их 62), кариотип которого включает 63 хромосомы. Реже у сук имеется всего лишь одна половая хромосома — Х0, что проявляется в синдроме Шерешевского-Тёрнера (с явлениями инфантилизма и отсутствием яичников). Кобели с набором XY:3A (половой индекс 0,33), вместо XY:2A (половой индекс равен 0,5), чрезмерно мускулистые, грубого сложения, с массивным костяком и называются сверхкобелями. Они также бесплодны. Иногда у кобелей бывает лишняя Y-хромосома, в этом случае у них, как и у мужчин с набором XYY, отмечается неоправданная и неуправляемая агрессия. НЕКОТОРЫЕ БИОХИМИЧЕСКИЕ АСПЕКТЫ В 1977 году М. Бёрнс [141], обсуждая мою книгу «Немецкая овчарка», опубликованную в 1976 году, высказала мнение, что я придерживаюсь несколько старомодного подхода к основам генетики. Это критическое замечание остается справедливым и по отношению к данной книге. С моей точки зрения, заводчики лучше усваивают генетику, объясняемую с точки зрения доминантных и рецессивных признаков, чем в свете более современных биохимических процессов. Однако замечание М. Бёрнс вполне справедливо, и для упрощенного объяснения действия генов я все-таки коротко поясню некоторые биохимические аспекты. До этого я привел вам в качестве аналогии нитку бисера для иллюстрации понятия генов и хромосом. В строгом смысле она некорректна, ибо гены — не изолированные объекты, а составляющие части хромосомы. Вся информация, необходимая для определения наблюдаемых нами в отдельной особи Один нуклеотид- Один нуклеотид Сахаро фосфат- Пары комплементарных " (Сахарофосфатный остов t оснований ный остов Одна полинуклео- н-связь Одна полинуклео- тидная цепь . . тидная цепь Рис. 14. ДНК (схематическое изображение развернутых цепей) А — аденин I I Сахарофосфатный '.ОСТОВ Две антипараллельные цепи 2 нм Рис. 13. Схематическое изображение структуры ДНК. На один полный оборот спирали приходится 10 пар оснований 3,4 км Основания- кольца Г — гуанин I Т — тимин ] Ц — цитозин I одно кольцо О фосфа°тДНЫе СВЯЗИ \ / Дезоксирибоза (сахар) признаков, содержится в структуре ДНК, из которой состоят гены. Сама ДНК — полимер, менее сложный по сравнению с белками, состоящий из сахара (дезоксирибозы), остатков молекул фосфорной кислоты и четырех азотистых оснований (пу- риновых — аденина, гуанина и пиримидиновых — цитозина и тимина). Молекула нуклеиновой кислоты представляет собой цепочку мономеров, специфических для этих кислот — нукле- отидов. Строение определенной молекулы нуклеиновой кислоты характеризуется определенной последовательностью нукле- отидов. Каждый нуклеотид состоит из трех разных элементов: азотистого основания, углеводного компонента и фосфорной кислоты. Для нуклеотидов ДНК характерна способность образовывать самые длинные молекулы из всех известных. Вероятно, каждый ген в хромосоме представляет собой участок ДНК, содержащий сотни или тысячи нуклеотидов. В 1953 году американский биохимик Джеймс Дьюи Уотсон и английский биофизик и генетик Фрэнсис Харри Комптон Крик выдвинули гипотезу, что молекула ДНК представляет собой двойную спираль, напоминающую лестницу со ступеньками из азотистых оснований, соединенных строго специфически (аденин с тимином, а гуанин с цитозином), перилами которой служат остатки фосфорной кислоты и пентозного сахара. Они же создали модель молекулы ДНК, а в 1962 году они вместе с английским биофизиком Морисом Уилкинсом впервые получили высококачественные рентгенограммы молекулы ДНК (см. рис. 13). Цепочки полинуклеотидов этой спирально завитой двойной молекулы расположены так, что сахар и фосфорные группы лежат снаружи молекул, а основания, соединенные между собой водородными связями, — внутри молекул (см. рис. 14). В отдельной цепочке из двадцати аминокислот хранится информация, отвечающая за синтез определенного белка, считываемая информационной, или матричной, РНК и переносимая транспортной РНК к рибосомной РНК. В определенных условиях двойная цепь ДНК разрывается на месте водородных связей и возле каждой цепи строится новая комплементарная цепь с помощью фермента полимеразы. В результате создаются две двойные цепи вместо одной. Так происходит процесс самокопирования ДНК: на ее позитивной цепочке синтезируется негативная, а на негативной — позитивная дополнительные цепочки. Две новые молекулы ДНК тождественны исходной и имеют такую же последовательность ступенек молекулярной лестницы. Способность к самокопированию — свойство только нуклеиновых кислот, которым не обладают даже белки. С помощью самокопирования осуществляется передача наследственных признаков, «записанных» на двойных спиралях ДНК. В одной молекуле нуклеиновой кислоты записано генетическое строение значительного количества различных белков. Это означает, что последовательность аминокислот в белке закодирована последовательностью нуклеотидов соответствующего участка молекулы нуклеиновой кислоты. Поскольку сведения о строении белков — самое главное, что должно быть передано потомкам при размножении, мы говорим о кодировании, то есть зашифровке наследственной информации — о биохимическом коде наследственности. Разнообразие фенотипов, наблюдаемое у животных, связано с различием в их ДНК. Когда дефект приписывается воздействию единичного гена, значит, произошло изменение на биохимическом уровне в единственной молекуле белка. Скорее всего, при этом обнаружится и специфическое изменение в основной цепочке ДНК. Так, из-за мутации вместо порядка AAA (три молекулы аденина) может возникнуть GAA, то есть на месте одной из молекул аденина появится молекула гуанина, что приведет к изменению кода, а значит, и наследственной информации. Чтобы доминантные гены вызвали биохимическую реакцию для фенотипического проявления признака, достаточно правильного сообщения, хранящегося только в одной из хромосом — только отцовской или материнской, тогда как для проявления рецессивного признака оно должно содержаться в обеих хромосомах — как в отцовской, так и в материнской. Подробно об этом написали в 1966 году Д.Ф. Паттерсон и У. Медуэй [710]. В своем труде за 1975 год [707] Д.Ф. Паттерсон приводит в качестве примера серповидноклеточную анемию (наследственная гемолитическая анемия) у человека, показав, что сложная клиническая картина этого тяжелого заболевания (периодический гемолиз, анемия, изменения со стороны костей и т. д.) связана с тем, что на определенной стадии биохимической реакции место валина занимает глутаминовая кислота, что, в свою очередь, приводит к замене аденина на определенном участке т-РНК тимином. Поскольку конечный продукт серии биохимических реакций зависит от правильного течения каждой из них, в самих сериях могут произойти изменения на разных этапах, способные привести к тому же самому результату. Так, независимо от причин, приводящих к гемофилии, заболевание проявляется в нарушении образования фактора VIII, ускоряющего свертывание крови. Ген, детерминирующий синтез этого фактора, находится в участке Х-хромосомы, не имеющей гомолога, и представлен двумя аллелями — доминантным нормальным и рецессивным мутантным. Нарушение любой из различных реакций в биохимической цепочке приведет к гемофилии, но на каждом этапе за них отвечают разные гены — одни из них сцеплены с полом, другие же — аутосомные. Наблюдая циклическую нейтропению (резкое уменьшение числа нейтрофилов в периферической крови с преобладанием юных форм) у длинношерстной колли, мы обнаруживаем у нее биохимические нарушения, не позволяющие нейтрофилам стать зрелыми, а также плейотропный эффект ослабления окраса шерсти, так называемый синдром серой колли, также связанный с изменением биохимических реакций, воздействующих на лейкоциты и формирование пигмента в волосах. Биохимическая генетика — самостоятельный раздел науки, хотя заводчиков, разумеется, больше интересуют не биохимические реакции, происходящие в организме животного, а более простые механизмы наследования. Если мы знаем, что конкретный дефект связан с дупликацией рецессивной аллели, то можем спланировать программу племенного разведения, и лежащие в основе всего этого биохимические процессы нас вовсе не интересуют. Как бы мы ни представили себе гены (в виде ДНК или нитки бисера), математика генетических коэффициентов от этого не изменится. |